 Eras geológicas.5 La era Cuaternaria o Cenozoica. Miocen Se denomina Mioceno, (del gr. "meion", menos, y "kainós", reciente), al cuarto período geológico de la Era Cenozoica. Comenzó hace 23,03 millones de años y terminó hace 5,332 millones de años.
Eras geológicas.5 La era Cuaternaria o Cenozoica. Miocen Se denomina Mioceno, (del gr. "meion", menos, y "kainós", reciente), al cuarto período geológico de la Era Cenozoica. Comenzó hace 23,03 millones de años y terminó hace 5,332 millones de años.En este período continuó la elevación de cordilleras como los Pirineos, los Alpes y el Himalaya.
La erosión favorecida por estas orogénesis originó sedimentos y depósitos de petróleo en zonas que eran cuencas marinas de poca profundidad. La temperatura era más baja que la actual y se originaron las masas de hielo en la Antártida.
Las plantas y los animales del Mioceno eran bastante modernos. El nombre de Mioceno significa "menos nuevo" aludiendo a que presenta un 18% menos de invertebrados marinos modernos que el Plioceno.
Los mamíferos y las aves estaban bien establecidos. Proliferaron las especies de mamíferos, entre ellos el rinoceronte, el gato, el camello y el caballo con las formas primitivas; entre estos están incluidos los grandes simios, que además de áfrica, vivían en Asia y en el sur de Europa.
En los mares, proliferaron ballenas y focas.
 Subdivisiones
SubdivisionesLa Comisión Internacional de Estratigrafía reconoce las siguientes edades/pisos del Mioceno:
* Mesiniense: 7,246 - 5,332 Ma
* Tortoniense: 11,608 - 7,246 Ma
* Serravalliense: 13,650 - 11,608 Ma
* Langhiense: 15,970 - 13,650 Ma
* Burdigaliense: 20,430 - 15,970 Ma
* Aquitaniense: 23,030 - 20,430 Ma
Estas subdivisiones se definen por la abundancia relativa de diferentes especies de nanofósiles calcáreos (algas unicelulares) y foraminíferos.
El Mioceno Superior, Medio y Inferior comprende cada uno de ellos dos subdivisiones.
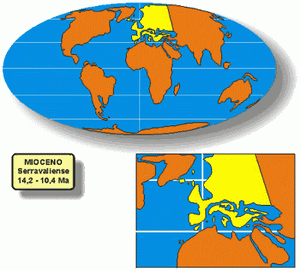 Paleogeografía
PaleogeografíaLos continentes continuaron la deriva hacia sus posiciones actuales. De las modernas características geológicas, sólo estaba ausente el puente terrestre entre Sudamérica y Norteamérica, aunque Sudamérica se estaba aproximando a la zona de subducción occidental en el Océano Pacífico, provocando tanto el aumento de los Andes como una extensión hacia el sur de la península mesoamericana.
La orogénesis continuó en el oeste de Norteamérica y Europa. Tanto los depósitos marinos como continentales del Mioceno son comunes en todo el mundo con abundantes afloramientos marinos cerca de las costas modernas.
Exposiciones continentales bien estudiadas se encuentran en las Grandes Llanuras de los Estados Unidos y en Argentina.
La India continuó chocando con Asia, aumentando el Himalaya. La vía Tetis continuó disminuyendo y se cerró cuando áfrica colisionó con Eurasia en la región turco-arábica hace entre 19 y 12 millones de años.
El posterior levantamiento de las montañas en la región del Mediterráneo occidental y un descenso global del nivel del mar se combinaron para causar una desecación temporal del Mar Mediterráneo (conocida como la crisis salina del Mesiniense) cerca del final del Mioceno.
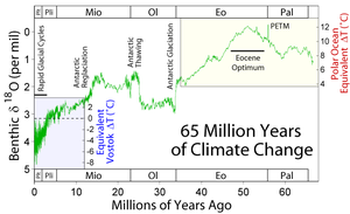 El clima cambiante del Mioceno
El clima cambiante del MiocenoToda la primera parte del Mioceno conserva el alto nivel de temperaturas alcanzado justo al final del Oligoceno.
Sin embargo, la transición del Oligoceno al Mioceno queda marcada por un corto episodio, llamado Mi-1, de intensa erosión en los continentes, que ocurre hace 23,7 millones de años, y que es apreciable a escala general del planeta.
Probablemente un enfriamiento y un aumento de la acumulación de hielo en la Antártida —especialmente en la Antártida Occidental— provocó una brusca bajada del nivel del mar, dejando las plataformas costeras en seco y expuestas a la erosión.
Todavía no existían mantos de hielo permanentes en el hemisferio norte, por lo que la fuerte bajada del nivel del mar debió deberse exclusivamente a la expansión del manto austral.
 Recientes investigaciones en sedimentos del mar de Ross indican que en esa transición entre el Oligoceno y el Mioceno se produjeron una serie de variaciones cíclicas en el volumen de hielo de la Antártida, ligadas a ciclos orbitales semejantes a los descritos por Milankovitch para el Cuaternario (Zachos, 2001; Naish, 2001).
Recientes investigaciones en sedimentos del mar de Ross indican que en esa transición entre el Oligoceno y el Mioceno se produjeron una serie de variaciones cíclicas en el volumen de hielo de la Antártida, ligadas a ciclos orbitales semejantes a los descritos por Milankovitch para el Cuaternario (Zachos, 2001; Naish, 2001).Optimo climático del Mioceno medio
Después de este corto pico inicial de frío las temperaturas se recuperan y se mantienen más altas que las del Oligoceno durante toda la primera mitad del Mioceno.
El volumen de los hielos de la Antártida vuelve a disminuir y el mar se eleva.
Las latitudes altas del hemisferio norte se calientan gradualmente y el paisaje de tundra de nuevo es sustituído por un paisaje de coníferas.
El nuevo máximo de calor se alcanza en el Mioceno medio, entre hace 17 Ma y 14,5 Ma (Mid-Miocene Climatic Optimum).
Los fósiles oceánicos y continentales indican que las temperaturas en las latitudes medias eran entonces 6°C superiores a las actuales.
 Mioceno final
Mioceno finalLa segunda parte del Mioceno, que comienza hace unos 14 Ma, es muy diferente a la primera. El estudio de la ratio Mg/Ca de foraminíferos planctónicos en el suroeste del Pacífico indica un enfriamiento brusco de 6°C a 7°C entre hace 14,2 y 13,8 Ma (Shevenell, 2004).
Las temperaturas se desploman y, sobre todo, el hielo continental aumenta en la Antártida.
Se atribuye este aumento del hielo en la Antártida a períodos de baja inclinación del eje terrestre. Esta baja oblicuidad provocaba menor intensidad en la insolación veraniega y además aumentaba el gradiente entre los polos y los trópicos, lo que motivaba una mayor transferencia meridiana de calor y de humedad hacia la Antártida.
Se produjo también una disminución del CO2, según se aprecia en el análisis isotópico del carbono (ratio 12C/13C) de sedimentos marinos.
Para algunos autores este momento representa así el paso de un clima “invernadero” al clima “nevera” en el que estamos metidos desde entonces (Holbourn, 2005).
Al final del Mioceno, entre hace unos 7 Ma y 5 Ma, un manto glacial acaba por cubrir, no sólo el continente austral, sino también toda Groenlandia.
Otro hecho climático importante, que acompaña este enfriamiento final, es el aumento de la aridez en vastas regiones de Asia y Africa, fenómeno que se agudizará en el Plioceno.
 Serravalliense-Aquitaniense
Serravalliense-AquitanienseEl clima se estaba calentando de nuevo. La acción combinada de las orogénesis y de determinados cambios en las corrientes oceánicas alteró las pautas de pluviosidad y de circulación atmosférica.
Un nuevo tipo de vegetación, el chaparral, empezó a formarse sobre todo en las vertientes continentales más áridas.
En los bosques subtropicales empezaron a aparecer claros herbáceos y en Sudamérica praderas.
Se abrieron rutas terrestres y las migraciones tuvieron un especial impacto sobre áfrica, a la que llegaron rumiantes, cerdos y verdaderos carnívoros procedentes de Eurasia.
En dirección contraria partieron cerdos hormigueros, proboscídeos y damanes. Los proboscídeos, rumiantes modernos y los carnívoros alcanzaron también Norteamérica.
Una característica esencial de este tiempo fue la aparición de pastos similares a la sabana en ambas Américas, por el enfriamiento global y la progresiva aridez del clima.
 Mesiniense-Tortoniense
Mesiniense-TortonienseEn el Asia oriental surgió una vegetación templada, aunque ni en áfrica, ni en Australia ni en Eurasia occidental apareció extensión alguna de auténticas praderas.
Los elefántidos ya se habían desarrollado, e incluso viajado desde áfrica hasta Eurasia.
Procedente de Norteamérica, el Hipparion alcanzó áfrica a través de Asia.
Las dos Américas estaban ya lo bastante próximas como para que los primeros «exploradores» se desplazaran entre ellas sobre restos de vegetación flotante.
La crisis salina del Mesiniense consistió en la desecación casi completa del Mediterráneo y tuvo lugar durante la era Mesiniense como consecuencia de la desconexión marina con el Océano Atlántico.
 La evaporación en el Mediterráneo supera la precipitación recogida por los ríos que en él drenan, por lo que la reducción del intercambio de agua con el Atlántico a través de la conexión del Rif hace 5,96 Ma (millones de años) y su clausura completa hace 5,59 Ma causó una rápida caída del nivel del Mar Mediterráneo al cancelarse el aporte de agua que actualmente proviene del Atlántico.
La evaporación en el Mediterráneo supera la precipitación recogida por los ríos que en él drenan, por lo que la reducción del intercambio de agua con el Atlántico a través de la conexión del Rif hace 5,96 Ma (millones de años) y su clausura completa hace 5,59 Ma causó una rápida caída del nivel del Mar Mediterráneo al cancelarse el aporte de agua que actualmente proviene del Atlántico.Esta evaporación produjo la deposición de grandes cantidades de sal en el fondo marino.
La crisis salina terminó con la inundación de enormes dimensiones del Mediterráneo por aguas Atlánticas hace unos 5.33 Ma, a través de un paso abierto en el actual Estrecho de Gibraltar.
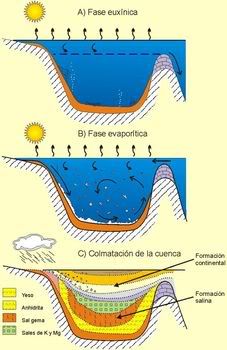
Durante cientos de miles de años, el paisaje del fondo del Mediterráneo, casi completamente desecado, debió asemejarse a una región semidesértica, con lagunas diseminadas de aguas salobres, hacia las que fluían los ríos a través de profundos cañones.
 Durante este período las aguas remanentes eran tan saladas que impedían la vida de la fauna marina.
Durante este período las aguas remanentes eran tan saladas que impedían la vida de la fauna marina.No se sabe con certeza el grado de desecación, pero la acumulación de sedimentos salinos llegó a alcanzar un espesor de hasta 2 y 3 km en algunas zonas.
Tales espesores son difíciles de explicar si no es porque se fuesen acumulando en sucesivas invasiones y evaporaciones de aguas saladas oceánicas.
Hoy día, la evaporación completa del actual Mediterráneo, si se cerrase Gibraltar, llevaría unos 1.000 años y en su fondo se formaría un sedimento de sales de 70 metros de espesor.
Por lo tanto, para acumular los 2 o 3 km de sedimentos del episodio mesiniense se habrían necesitado 30 o 40 ciclos de llenado y secado de la cuenca.
 En la Península Ibérica, se configuran las dos grandes cuencas de la meseta (Duero y Tajo), que habían iniciado su estructuración en el período anterior.
En la Península Ibérica, se configuran las dos grandes cuencas de la meseta (Duero y Tajo), que habían iniciado su estructuración en el período anterior.Numerosos retazos de pequeñas cuencas, como la del norte de Extremadura, parecen prolongar ambas fosas hacia Portugal, convergiendo en la cuenca portuguesa del Tajo que se abre en este período al océano Atlántico.
En el Aquitanense tiene lugar un intercambio biótico con Europa del noreste y con áfrica, aunque se mantiene alta la tasa de endemismo.
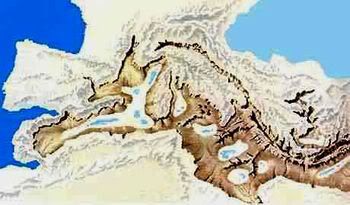
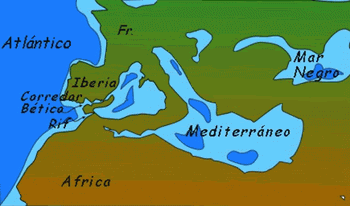
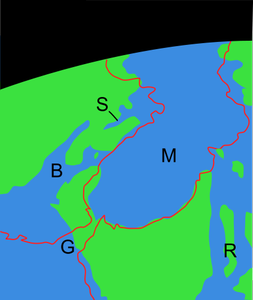
Junto a estas líneas, una reconstrucción paleogeográfica del sur de la Península Ibérica durante el Mioceno previo a la formación del Estrecho de Gibraltar.
La línea roja indica la costa actual. S=Cuenca de Sorbas, Almería, B=Corredor bético, R=Corredor del Rift. G=Estrecho de Gibraltar, M=Mar Mediterráneo.
 El Mesiniense finalizó justo en la frontera entre el Mioceno y el Plioceno, hace unos 5,4 millones de años. Entonces otro cataclismo orogénico en el extremo occidental del Mediterráneo volvió a abrir la comunicación con el Atlántico.
El Mesiniense finalizó justo en la frontera entre el Mioceno y el Plioceno, hace unos 5,4 millones de años. Entonces otro cataclismo orogénico en el extremo occidental del Mediterráneo volvió a abrir la comunicación con el Atlántico.Esta vez, por Gibraltar. Y desde entonces el Mediterráneo se encuentra en equilibrio gracias al agua superficial que entra desde el Atlántico, que compensa la pérdida por evaporación que sufre la cuenca (mayor que la precipitación y el aporte de los ríos) y también la pérdida de agua muy salada que se escapa al Atlántico en dirección contraria por las profundidades del Estrecho.
Las posibles causas del cierre de la conexión con el Atlántico en la zona del Paso del Rif (conexión anterior de la Crisis Mesiniense):
* Un levantamiento tectónico de la zona del Corredor del Rif (Arco de Gibraltar). Se han propuesto tres posibles contextos tectónicos para la región geológica del Arco de Gibraltar:
* Retirada de la placa subducente (slab rollback) durante una subducción. Se han propuesto distintas orientaciones para la orientación de dicha subducción.
* Delaminación mantélica y hundimiento de una capa de la litosfera terrestre en la región.
* Colapso gravitacional y desgarre de la raiz litosférica de un orógeno anterior al Messiniense.
* Un descenso global del nivel del mar que dejase emergido un istmo entre Europa y áfrica.
El nivel del mar era entre 6 y 23 m más bajo que actualmente hace 5.96 Ma y entre 11 y 19 m hace 5.59 Ma (Miller et al., 2005, Science).
 El levantamiento isostático de la zona del Paso del Rif en respuesta a la eliminación del peso del agua que reposaba sobre la litosfera fue de entre decenas y centenares de metros, contribuyendo así a impedir una pronta reinundación del Mediterráneo.
El levantamiento isostático de la zona del Paso del Rif en respuesta a la eliminación del peso del agua que reposaba sobre la litosfera fue de entre decenas y centenares de metros, contribuyendo así a impedir una pronta reinundación del Mediterráneo.En esta época destaca la peculiaridad y el gran desarrollo que adquiere la sedimentación, la actividad geodinámica y los cambios biogeográficos, especialmente en el Mediterráneo occidental.
El depósito de grandes espesores de evaporitas, actualmente bajo el mar, ha llevado a la interpretación de la probable y repetida desecación del Mediterráneo en el Mesiniense, lo que incentivó notablemente las investigaciones.
En cada una de las desecaciones el Mediterráneo se convertía en una enorme cuenca desértica que, en ciertos puntos, llegaba a alcanzar la cota de 5000 metros por debajo del nivel del mar.
Después de cada episodio de desecación, la cuenca era inundada de nuevo por la entrada torrencial de aguas del Paratetis y Atlántico por el Estrecho de Gibraltar.
Al final del Mesiniense, numerosos inmigrantes testimonian que un activo intercambio faunístico tuvo lugar en el área entre Asia, áfrica y Europa durante la regresión de la cuenca mediterránea.
Recientemente la desecación del Mediterráneo ha sido cuestionada por algunos investigadores.
 Paleobiología
PaleobiologíaLas Globigerinas, seriamente diezmadas en la extinción eoceánica, se expanden de nuevo en el Mioceno y constituyen unos excelentes fósiles guía en los medios marinos.
Las ddiatomeasd de agua dulce (Pennales), comprenden 2000 especies alcanzando una posición ecológica similar a la actual como productores primarios de agua dulce.
Los cambios climáticos ejercieron una influencia profunda sobre las biotas terrestres. El mejor indicador de los cambios climáticos son los cambios evolutivos y de distribución geográfica de las angiospermas.
La gran abundancia de plantas herbáceas en este período permitiría llamarlo Edad de las Hierbas.
El éxito de las plantas herbáceas es el resultado del deterioro climático generalizado que tuvo lugar durante el Oligoceno y Mioceno.
El enfriamiento del clima y unas condiciones más secas, ocasionaron la regresión de los bosques y la presencia de plantas herbáceas que prefieren los hábitats abiertos y pueden resistir bajas precipitaciones.
 Criaturas consideradas «menores» experimentaron un gran éxito: paseriformes, ranas, ratas, ratones, serpientes, etc.
Criaturas consideradas «menores» experimentaron un gran éxito: paseriformes, ranas, ratas, ratones, serpientes, etc.Las familias de rinocerontes y caballos disminuyeron después del Mioceno medio en una continuación del decaimiento general de los ungulados de dedos impares.
Mientras tanto, Cervidae y Bovidae se diversificaron, aunque el número de especies de estas familias ha disminuido desde entonces.
De forma similar, muchos tipos de elefantes, incluyendo los que tenían largas trompas, experimentaron un gran éxito durante el intervalo Mioceno-Plioceno, para luego declinar (hoy sólo existen dos especies). Aparecen las familias de las hienas y osos.
 Amphicyonidae es una familia de fósiles (conocidos como oso-perro) pertenecientes al orden Carnivora.
Amphicyonidae es una familia de fósiles (conocidos como oso-perro) pertenecientes al orden Carnivora.Tradicionalmente se creía que esta familia estaba emparentada con la familia Ursidae. Sin embargo hay evidencias que apunta a un parentesco con la familia Canidae. La talla era similar a la del actual oso negro.
Amphicyon, conocido también como oso-perro. De constitución robusta, podía alcanzar los 2 metros de largo. Tenía una cola larga y pesada, cuello grueso, extremidades robustas y dientes similares a los de los lobos.
Se han encontrado restos fósiles de Amphicyon en Norteamérica y Europa. Durante el Mioceno inferior, una gran cantidad de amphicyonidos migraron desde Eurasia hasta Norteamérica.
Esos taxones pertenecen a la subfamilia Amphicyoninae.
El primero en aparecer fue Ysengrinia, seguido de Cynelos y de Amphicyon. Este flujo de amphicyonidos, acompañados por otros ungulados y pequeños mamíferos, indican un gran intervalo (desde 23 hasta 16.5 Ma) de intercambio de fauna entre Eurasia y Norteamérica en el Mioceno inferior.
 Megatherium americanum es una especie de mamífero placentario del orden Pilosa. Fue un perezoso terrestre gigante que habitó en America hasta hace 9.000 años y que fue descrito en 1769 por Georges Cuvier.
Megatherium americanum es una especie de mamífero placentario del orden Pilosa. Fue un perezoso terrestre gigante que habitó en America hasta hace 9.000 años y que fue descrito en 1769 por Georges Cuvier.Medía 5 metros desde la cabeza hasta la cola y 2 m. desde el suelo hasta el lomo, se sabe que estaba cubierto de pelo rojizo y que su piel contenía pequeños huesecillos los cuales la hacían tan dura como un chaleco antibalas.
Era terrestre, lo que se deduce por su tamaño y de hábitos solitarios. Comía plantas apoyado en sus patas traseras aunque podía usar las cuatro apoyándose en los nudillos como los gorilas.
 Sinomastodon ("mastodonte chino") es un género extinto de proboscidios que habitaron entre el Mioceno superior y el Pleistoceno inferior en algunos países del sureste asiático como China, Japón, e Indonesia.
Sinomastodon ("mastodonte chino") es un género extinto de proboscidios que habitaron entre el Mioceno superior y el Pleistoceno inferior en algunos países del sureste asiático como China, Japón, e Indonesia.Este animal era muy parecido a los elefantes actuales, con un tamaño que oscilaba entre los 3,6 y 5,3 metros.
Se conocen varios ejemplares chinos, de los cuales el más conocido es el S. hanjiangensis del Mioceno superior y el Plioceno inferior sobre la provincia de Shanxi.
Se le conoce a partir del esqueleto incompleto de un ejemplar adulto de 5,3 metros, el cual incluía dientes, maxilares, colmillos, y otros materiales.
La especie japonesa S. sendaicus, descrita en 1924 a partir de partes de dentición halladas en yacimientos del Plioceno, ha sido clasificada dentro del género, así como la especie S. bumiajuensis (antiguamente Tetralophodon) del Plioceno superior de la isla de Java.
 Purussaurus es un género extinto de cocodrilianos. Se trata de un caimán gigante que vivió en Suramérica, hace 20 millones de años.
Purussaurus es un género extinto de cocodrilianos. Se trata de un caimán gigante que vivió en Suramérica, hace 20 millones de años.Es conocido únicamente por materiales del cráneo hallados en la Amazonía peruana.
El cráneo tiene cerca de 1,5 metros de largo, por lo que algunos paleontólogos estiman que el cuerpo entero debió medir cerca de 15 m, lo que significaría que el Purussaurus es uno de los mayores cocodrilos que se conozcan.
 En los restos de anfibios del yacimiento de azufre de Libros (Teruel), los cuales gracias a las condiciones existentes cuando se formó el yacimiento poseen un estado de conservación excelente, encontrándose los animales completos y con el contorno del cuerpo conservado a modo de impronta en las margas pizarrosas del yacimiento del Mioceno.
En los restos de anfibios del yacimiento de azufre de Libros (Teruel), los cuales gracias a las condiciones existentes cuando se formó el yacimiento poseen un estado de conservación excelente, encontrándose los animales completos y con el contorno del cuerpo conservado a modo de impronta en las margas pizarrosas del yacimiento del Mioceno.Esta conservación, muy rara en fósiles de vertebrados, se debe al ambiente reductor de las aguas donde caen los animales muertos, que impedían la destrucción de los tejidos blandos por las bacterias de la putrefacción, dando tiempo a que los sedimentos cubriesen los cuerpos de estos animales, procesos posteriores de compactación y de diagénesis formaron estos fósiles.
Longinos Navás (S.J.) eminente naturalista y científico de la transición del siglo XIX al XX e interesado en ramas muy diversas de la Paleontología y Biología, definió varias especies del género Rana en el yacimiento mioceno de Libros (Teruel), dentro de las que destacan Rana pueyoi y Rana quellenbergi.
El grupo de los primates del Viejo Mundo se amplió, con la aparición de los monos cercopitécidos (en los que hoy se encuadran macacos y babuinos).
Los primeros homínidos (Sahelanthropus, Ardipithecus) empezaron a diversificarse en el este de áfrica.
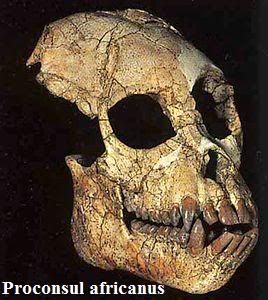 El Mioceno Superior se cree que es cuando se produce una radiación importante de los simios, de los existentes en el Mioceno medio y el surgimiento de nuevos (sobre todo en Europa y Asia).
El Mioceno Superior se cree que es cuando se produce una radiación importante de los simios, de los existentes en el Mioceno medio y el surgimiento de nuevos (sobre todo en Europa y Asia).Según los dato moleculares es cuando se produce la separación de los linajes de los grandes simios asiáticos y africanos (orangután, gorila y chimpancé). Pero por desgracia no tenemos los mismos datos en el registro fósil.
Entre 17-12 m.a. se han encontrado restos de homínidos en áfrica, Europa y asía, pudiéndose agrupar en varias Subfamilias, dentro de la familia Afropithecinae (aún en discusión).
Todavía aún no está claro si las especies o géneros encontrados pertenecen a una u otra subfamilia.
Se cree que estas subfamilias evolucionan a partir del genero Procónsul en áfrica, principalmente en Kenia.
Debido a los cambios geológicos que tuvieron lugar en aquellos momentos, favoreció que estas subfamilias, pudiera colonizar nuevos nichos ecológicos fuera de áfrica, encontrándose restos de estas, como el genero,Heliopithecus. Que aparece hace 17 m.a, Ab Dabtiyah, centro de Arabia Saudita, y puede que en Turquía.
Entre el Mioceno Medio-Superior ,13 y 8 m.a., aparecen los siguientes géneros más importantes:
 En Europa:
En Europa:-Dryopithecus (aparece en el a finales del Mioceno medio).
-Oreopithecus.
-Ouranopithecus.
Asia:
-Ankaraithecus.
-Sivapithecus.
-Lufengpithecus.
-Otavipithecus.
-Gigantopithecus.
Todos muestran adaptaciones propias de la vida en bosques tropicales.
El Dryopithecus, se encontró por primera vez en Francia, y a partir de su fauna y se le atribuyó unos 13 millones de años, por lo que habría que ubicarlo a mediados del Mioceno Medio.
 En los océanos, aparecieron los tiburones modernos, en particular el gran Megalodon y evolucionan los cetáceos, como delfines, ballenas y marsopas.
En los océanos, aparecieron los tiburones modernos, en particular el gran Megalodon y evolucionan los cetáceos, como delfines, ballenas y marsopas.Alrededor de 23 millones de años atrás los Kentriodon, unos "delfines" de la antigüedad aparecieron con sistema de ecolocación. Al parecer se hicieron bastante comunes 14 a 16 milllones de años, hasta que hace unos 10 a 12 millones de años aparecieron los primeros delfines "modernos".
Cetotherium es un género extinto de cetáceos que se asemejaba bastante a las actuales especies de cetáceos marinos.
 Vivió en el Mioceno, hace unos 15 millones de años aproximadamente. En su tiempo fue presa del Megalodon y de otros predadores marinos. Los cetoterios alcanzaban una longitud de unos 12 metros.
Vivió en el Mioceno, hace unos 15 millones de años aproximadamente. En su tiempo fue presa del Megalodon y de otros predadores marinos. Los cetoterios alcanzaban una longitud de unos 12 metros.Los actuales misticetos son animales que se alimentan mediante el filtrado del agua con sus "barbas".
Los primeros individuos con esta característica aparecieron a mediados del Mioceno.
Esta y otras características aparecieron como resultado de cambios físicos y medioambientales en los océanos. Un cambio a gran escala en las corrientes y en la temperatura del agua pudo haber propiciado el despegue de los modernos misticetos y el declive de las especies arcaicas.
 El Carcharodon megalodon, llamado también megalodonte, tiburón megadiente y tiburón blanco gigante, fue un tiburón gigante que vivió probablemente entre hace 20 y 1,6 millones de años, aunque se ha llegado a sugerir fechas de extinción más tardías e incluso que pudiera sobrevivir hoy en día.
El Carcharodon megalodon, llamado también megalodonte, tiburón megadiente y tiburón blanco gigante, fue un tiburón gigante que vivió probablemente entre hace 20 y 1,6 millones de años, aunque se ha llegado a sugerir fechas de extinción más tardías e incluso que pudiera sobrevivir hoy en día. El megalodon se conoce únicamente por numerosos dientes fósiles y algunas vértebras y esqueletos parciales en el Caribe.
El megalodon se conoce únicamente por numerosos dientes fósiles y algunas vértebras y esqueletos parciales en el Caribe.Los dientes son en muchos aspectos similares a los del tiburón blanco actual (Carcharodon carcharias), pero con un tamaño que puede superar los 17,5 centímetros de largo, por lo que se suele considerar la existencia de un estrecho parentesco (y probablemente, también parecido aspecto y conducta) en ambas especies.
Sin embargo, algunos investigadores opinan que las similitudes entre los dientes de ambos animales son producto de un proceso de evolución convergente.
Las estimaciones más sensatas del tamaño de esta criatura oscilan entre los 12 y 15 metros. Las reconstrucciones previas con longitudes que podían llegar a los 30 metros se consideran por lo general como poco precisas.
Existe la teoría de que los megalodones adultos se alimentaban de ballenas y se extinguieron cuando los mares polares se volvieron demasiado fríos para los tiburones, permitiendo que las ballenas estuviesen a salvo de ellos durante el verano.